Start with a case in which behaviour reading is clearly involved.
I take Byrne’s study
to demonstrate that chimps are capable of sophisticated behaviour reading.
But how might they represent behaviours?
The procedure for preparing a nettle to eat while avoiding contact with its stings is shown in
\vref{fig:byrne_2003_fig1}. It involves multiple steps. Some steps may be repeated varying numbers of
times, and not all steps occur in every case. The fact that gorillas can learn this and other
procedures for acquiring and preparing food by observing others’ behaviour suggests that they have
sophisticated behaviour reading abilities \citep[p.~513]{Byrne:2003wx}. If we understood the nature
of these behaviour reading abilities and their limits, we might be better able to understand their
abilities to track mental states too.
Byrne 2003, figure 1
‘great apes [are] able to acquire complex and elaborate local traditions of food
acquisition, some of them involving tool use’ \citep[p~513]{Byrne:2003wx}
So even quite sophisticated behaviour reading is possible without any
reliance on communication by language.
We can therefore think of behaviour reading as foundational for any kind
of radical interpretation.
[background]
‘Nettles, Laportea alatipes, are an important food of mountain gorillas in Rwanda (Watts 1984),
rich in protein and low in secondary compounds and structural carbohydrate (Waterman et al. 1983).
Unfortunately for the gorillas, this plant is 'defended' by powerful stinging hairs,
especially dense on the stem, petioles and leaf-edges.
All gorillas in the local population process nettles in broadly the same way,
a technique that minimizes contact of sting- ing hairs with their hands and
lips (Byrne & Byrne 1991; figure 1). A series of small transformations is
made to plant material: stripping leaves off stems, accumulating
larger bundles of leaves, detachment of petioles, picking out unwanted debris,
and finally folding a package of leaf blades within a single leaf before
ingestion. The means by which each small change is made are idiosyncratic and
variable with context (Byrne & Byrne 1993), thus presum- ably best learned
by individual experience. However, the overall sequence of five discrete
stages in the process is standardized and appears to be essential for efficiency
(Byrne et al. 2001a).’ \citep[pp.~531--2]{Byrne:2003wx}
‘Like other complex feeding tasks in great apes, preparing nettles is a hierarchically
organized skill, showing considerable flexibility: stages that are occasionally
unnecessary are omitted, and sections of the process (of one or several ordered stages)
are often repeated iteratively to a criterion apparently based on an adequate size of
food bundle (Byrne & Russon 1998).’ \citep[pp.~532]{Byrne:2003wx}
Byrne 2003, figure 1
Recall Byrne’s study.
To understand this action, the Teleological Stance is insufficient.
Why?
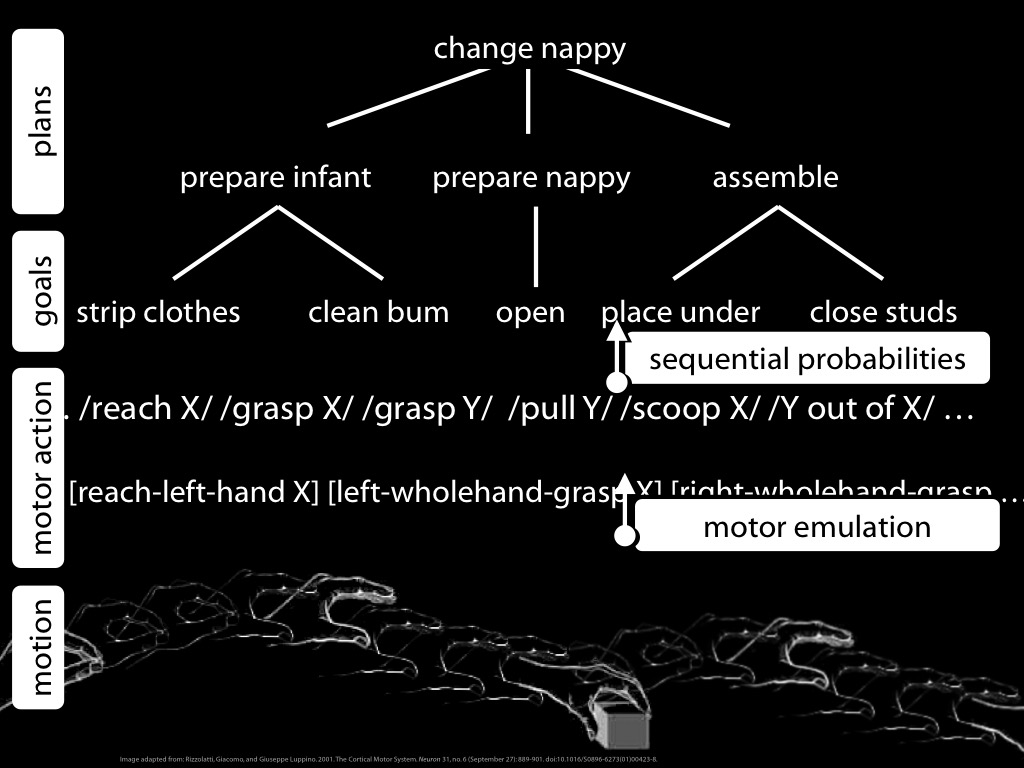
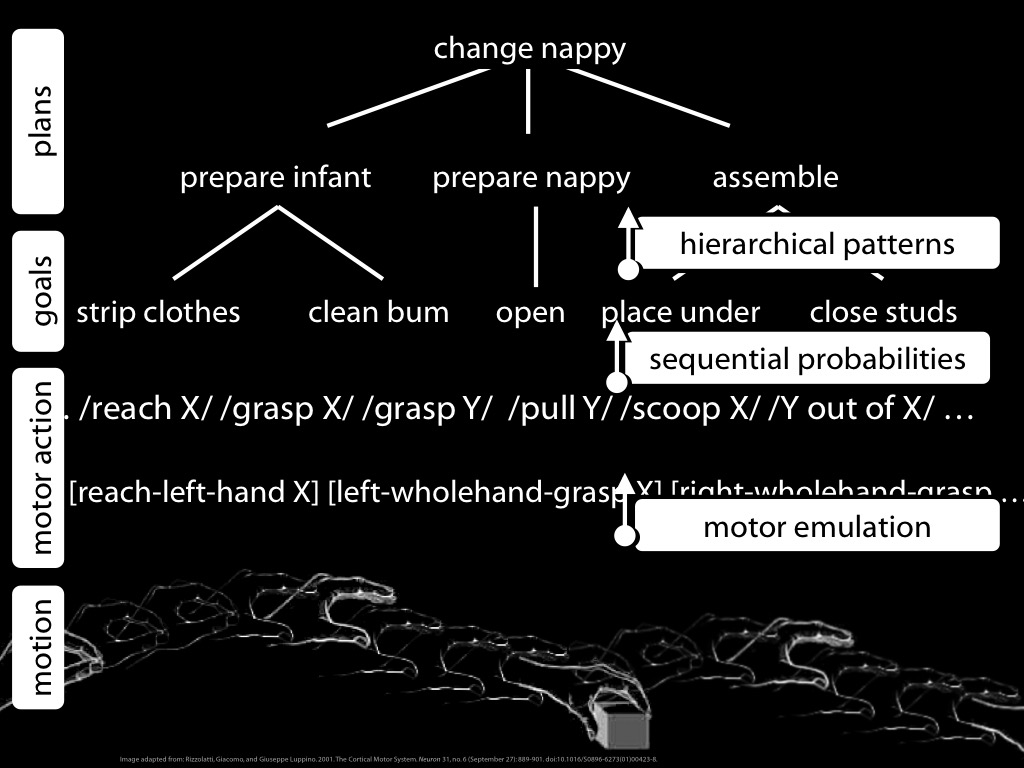
Let me recap the four ingredients with a diagram ....





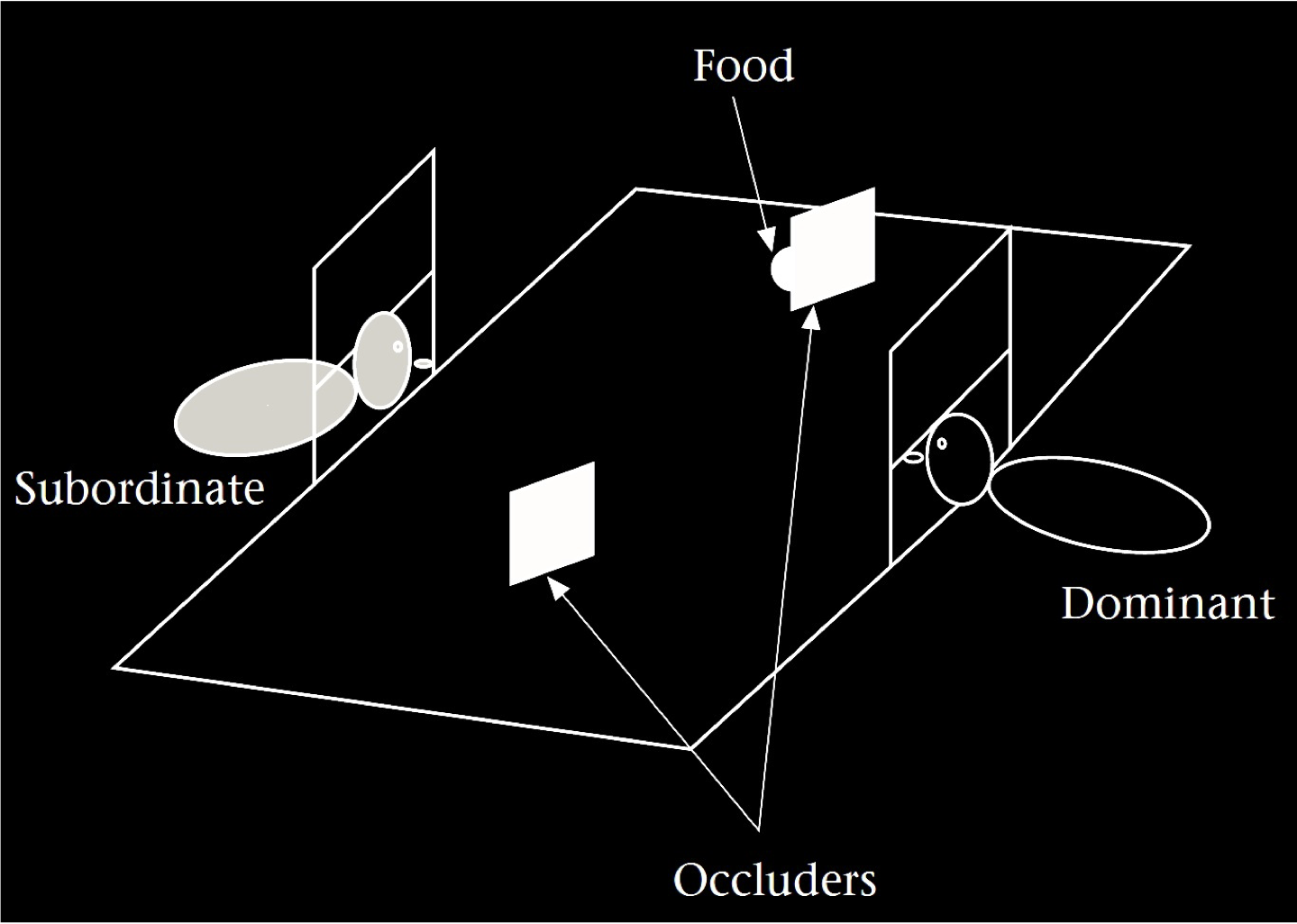
Hare et al (2001, figure 1)
Although not designed to test such limits, some existing experimental designs involve features which plausibly exclude explaining subjects’ performance in terms of pure behaviour reading only.
To illustrate, consider the sequence of events in the ‘misinformed’ condition of \citet[][Experiment 1]{Hare:2001ph}.
A competitor observes food being placed [$A$], the competitor’s access is blocked [$B$], stuff happens [$X^N$], food is moved [$C$], more stuff happens [$Y^N$], and the competitor’s access is restored [$D$].
Finding evidence that chimpanzees can learn to identify patterns of this form [$ABX^NCY^ND$] and use them to predict the conspecifics’ behaviours would represent a major discovery.
‘Before the subordinate’s door was raised, thus allowing her to enter the middle cage, we closed the dominant’s door, regard- less of whether it had been open before, and it remained closed until the subordinate’s hand first touched the floor of the middle cage.’ Hare et al (2001, p. 142)